246|Evangelina Palacios
Estimulación por la insulina de la biosíntesis de novo. La insulina (Figura 5)
activa a las enzimas que intervienen en la lipogénesis y con ello incrementa, el
reservorio de AG en los adipocitos. La vía lipogénica es activa en los adipocitos
aunque en menor proporción que en el hígado.
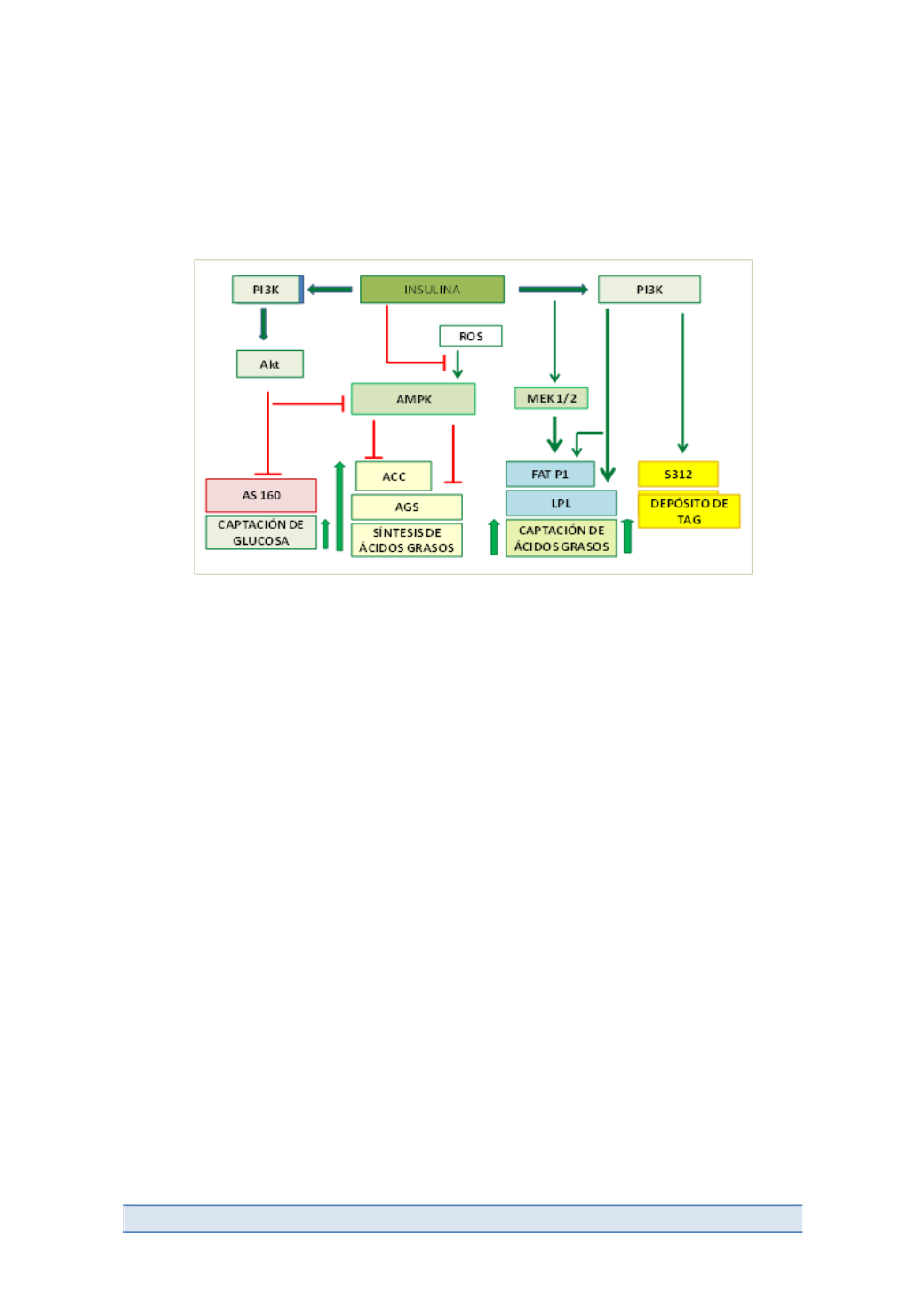
Figura 5.
La señalización por insulina estimula la captación rápida de glucosa y su transporte, la
biosíntesis “de novo” de los ácidos grasos así como la captación de mismos (con intervención de la
LPL) y su esterificación para formar TAG.
En adipocitos aislados, se ha demostrado la inactivación, por fosforilación,
de la acetil-‐CoA carboxilasa (ACC) enzima limitante de velocidad en la síntesis de
AG. En la reacción catalizada por ACC se consume acetil CoA y se produce malonil
CoA, que es sustrato necesario para biosíntesis citosólica de ácidos grasos
(palmitato) .La fosforilación de ACC, se lleva cabo por la AMPK. Esta enzima
(quinasa dependiente de AMP) se inactiva por fosforilación, modificación
covalente, que puede ser llevada a cabo por la Akt. Así, la señalizacion por insulina
a través de la PI-‐3K/Akt (Figura 5) conduce a fosforilación) de AMPK. Esta quinasa,
inactiva, está incapacitada para fosforilar e inactivar a la ACC que permaneciendo
en su estado no fosforilado y activo, proporciona el sustrato lipogénico, malonil
CoA. En definitiva, la insulina favorece la biosintesis citosólica de ácidos grasos.
Por otro lado, la señalización por insulina bloquea la activación de AMPK por las
especies reactivas de oxígeno (ROS) (Figura 5) y mediante esa vía conduce,
también, a la incapacitación de la quinasa para fosforilar e inactivar a la ácido


