ARTÍCULO |
Estudio de la actividad neuroprotectora de diterpenos aislados del género Sideritis
Elena Mª González Burgos, Mª Emilia Carretero Accame, Mª Pilar Gómez-Serranillos Cuadrado*
Departamento de Farmacología, Facultad de Farmacia, Universidad Complutense de Madrid, Ciudad Universitaria 28040– Madrid, España
*e-mail: egonzalezburgos@hotmail.com
An. Real Acad. Farm. Vol. 80, Nº 2 (2014), pag. 362-376.
Resumen de la Tesis Doctoral premiada con el premio “Antonio Doadrio López” en el Concurso Científico 2013 de la Real Academia Nacional de Farmacia.
resumen
El estrés oxidativo, definido como un desequilibrio entre la producción de especies reactivas de oxígeno (ROS) y la capacidad del sistema defensivo del organismo (enzimático y no enzimático) para hacer frente a las mismas, está implicado en la fisiopatología de varias enfermedades neurodegenerativas incluidas la enfermedad de Alzheimer, la enfermedad de Parkinson, la enfermedad de Huntington y la Esclerosis Lateral Amiotrófica. En este contexto, la búsqueda de compuestos antioxidantes que permitan disminuir o neutralizar el daño oxidativo constituye un importante campo de investigación. En este trabajo de Tesis Doctoral se ha evaluado la actividad neuroprotectora frente a estrés oxidativo de los diterpenos andalusol, conchitriol, foliol, lagascatriol, linearol y sidol aislados de especies del género Sideritis. |
Palabras clave: diterpenos; Sideritis; neuroprotección.
abstract
Oxidative stress, defined as a disturbance in the equilibrium between reactive oxygen species (ROS) production and the capacity of the defense system of the organism (enzymatic and non-enzymatic) to protect against them, is involved in the physiopathology of several neurodegenerative diseases including Alzheimer's disease, Parkinson's disease, Huntington's disease and Amyotrophic Lateral Sclerosis. In this context, the search of antioxidant compounds is an interesting research field. In this Doctoral Thesis, it has been evaluated the neuroprotective activity against oxidative stress of the diterpenes andalusol, conchitriol, foliol, lagascatriol, linearol and sidol isolated from Sideritis spp. |
Keywords: diterpenes; Sideritis; neuroprotection.
1. INTRODUCcióN
Las enfermedades neurodegenerativas como la enfermedad de Alzheimer, la enfermedad de Parkinson, la Esclerosis Lateral Amiotrófica y la enfermedad de Huntington son procesos crónicos caracterizados por la pérdida progresiva de las neuronas y la consiguiente alteración de las funciones motora, sensorial y cognitiva. Estas enfermedades constituyen un problema de salud pública a nivel mundial, que crece día a día debido al vertiginoso envejecimiento de la población. La investigación básica y clínica aúna esfuerzos para conocer mejor la fisiopatología de estas enfermedades así como para desarrollar nuevas estrategias terapéuticas tanto para paliar como para frenar la progresión de las mismas (1, 2).
Durante las últimas décadas, numerosos estudios experimentales in vitro e in vivo, así como estudios post mortem en tejidos cerebrales humanos, apoyan la implicación del estrés oxidativo en la patogenia de diversas enfermedades neurodegenerativas, incluidas las anteriormente mencionadas (3, 4). El estrés oxidativo, definido como una alteración de la homeostasis óxido-reducción intracelular consecuencia de una excesiva producción de ROS y/o de una deficiencia en los mecanismos antioxidantes endógenos de tipo enzimático y/o no enzimático, puede conllevar un daño celular irreversible e irreparable por mecanismos como son la peroxidación lipídica en las membranas celulares, la modificación de las bases de los ácidos nucleicos, la inactivación de las enzimas y la oxidación de las proteínas; incluso conducir a la muerte celular, principalmente por apoptosis (5, 6).
El uso de antioxidantes exógenos se presenta como una de las estrategias terapéuticas más prometedoras para disminuir o neutralizar el daño oxidativo ocasionado por las especies reactivas de oxígeno (ROS) (7). Los mecanismos de acción descritos que definen la capacidad antioxidante de un compuesto son diversos: a) interacción directa con especies reactivas o actividad "scavenger" de radicales libres, vía "transferencia de un solo electrón" (SET) o vía "transferencia de un átomo de hidrógeno" (HAT), b) quelación de metales de transición (hierro, cobre) que pueden iniciar reacciones de oxidación, c) inducción o activación de las enzimas antioxidantes (catalasa, superóxido dismutasa, glutation peroxidasa, glutation reductasa, hemooxigenasa-1, entre otras), lo que se traduce en un mayor equilibrio redox celular y d) inhibición de la actividad, expresión o síntesis de enzimas prooxidantes como NADPH-oxidasa, mieloperoxidasa, óxido nítrico sintasa y xantina-oxidasa, implicadas en la generación de especies reactivas (8). En este contexto, los productos naturales constituyen una fuente inagotable de principios activos con potencial antioxidante. De hecho, recientes investigaciones han demostrado la capacidad antioxidante de diversos compuestos de origen natural, siendo entre ellos relevantes el grupo de los diterpenos (9).
El género Sideritis, perteneciente a la familia de las Lamiaceae, comprende aproximadamente 150 especies distribuidas por la vertiente mediterránea europea, africana y parte de la asiática, extendiéndose desde las islas Canarias y Madeira hasta el Cáucaso (10). Distintas especies de Sideritis se han usado tradicionalmente para preparar infusiones, como agentes saborizantes así como con fines terapéuticos por sus propiedades antiinflamatorias, antioxidantes, antiulcerosas, digestivas, antimicrobianas, vulnerarias, analgésicas y carminativas (11). Más de 160 diterpenos, incluidos en al menos 9 grupos de estructuras diferentes, han sido identificados y aislados de las partes aéreas de Sideritis spp. (12). Los compuestos sideridiol y siderol fueron los primeros diterpenos aislados y su estructura se elucidó durante la segunda mitad del siglo XX (13, 14).
Este trabajo de Tesis Doctoral se ha planteado como el estudio de la actividad neuroprotectora, frente al estrés oxidativo, de los diterpenos andalusol, conchitriol, lagascatriol, foliol, linearol y sidol aislados de especies del género Sideritis y su posible aplicación en aquellos procesos neurodegenerativos que cursen con daño celular.
2. material y métodos
2.1. Diterpenos
El diterpeno de tipo labdanoandalusol se aisló de la especie Sideritis arborescens Salzm. (15). Tanto el diterpeno de tipo beyerano, conchitriol, como el de tipo rosano, lagascatriol, se aislaron de Sideritis angustifolia Lag. (16-18). Los diterpenos derivados del kaurano, foliol, linearol y sidol se obtuvieron de Sideritis linearifolia Lam. (18).
2.2. Líneas celulares
2.2.1. Líneas celulares utilizadas en los estudios de estrés oxidativo y disfunción mitocondrial. Tratamiento celular.
Línea celular PC12 (Neuronas)
La línea celular PC12 se adquirió de la colección de cepas American Type Culture Collection (ATCC). El medio de cultivo utilizado es Dulbecco’s Modified Eagle’s Medium (DMEM) (Gibco, Invitrogen Corporation) suplementado con 0,5% de gentamicina (Gibco, Invitrogen Corporation) y 0,2% de piruvato (Sigma-Aldrich). Las células se mantienen a 37ºC en atmósfera de aire al 95% y de CO2 al 5%.
Línea celular U373-MG (Astrocitos)
La línea celular U373-MG se adquirió desde Sigma-Aldrich procedente de la Colección Europea de Cultivos Celulares Animales (ECACC). El medio de cultivo empleado es Dulbecco’sModifiedEagle’s Medium (DMEM) (Gibco, Invitrogen Corporation) suplementado con 0,5% de gentamicina (Gibco, Invitrogen Corporation) y suero fetal bovino (FBS) (Gibco, Invitrogen Corporation).Las células se mantienen en condiciones de temperatura y atmósfera húmeda de 37ºC, 95% de aire y 5% de CO2.
Tratamiento celular
Las células U373-MG y PC12 se tratan con los diterpenos objeto de estudio (diferentes concentraciones según el propio desarrollo de la Tesis) durante 24 h, y posteriormente con H2O2 (1 mM para las células U373-MG y 0,1 mM para las células PC12) durante 30 minutos.
2.2.2. Líneas celulares utilizadas en los estudios de paso a través de Barrera Hemato Encefálica (BHE)
Línea celular RBE4
Las células RBE4 fueron cedidas por el Dr. Pierre Olivier Couraud del Instituto Cochin del Instituto Nacional de la Salud y de la Investigación Médica (INSERM) de París (Francia).Las células RBE4 crecen en frascos de cultivo tapizados durante al menos 3 horas con 3 mL de una solución de colágeno obtenida a partir del tendón de cola de rata. Las células se mantienen en una atmósfera de 95% de aire y 5% de CO2, saturada de humedad a 37ºC. El medio de cultivo empleado es Hams F-10/α-Minimum Essential Medium (α-MEM) (1:1) (Gibco, Invitrogen Corporation), suplementado con Glutamax (Sigma-Aldrich), 10% de suero fetal de ternero (Gibco, Invitrogen Corporation), geneticina (300 μg/mL) (Sigma-Aldrich) y factor básico de crecimiento de fibroblastos (bFGF, 1 ng/mL) (Boehringer).
Línea celular ECV304/C6
La línea celular ECV304 corresponde a células endoteliales de cordón umbilical humano. Esta línea celular se obtuvo de la Colección Europea de Cultivos Celulares Animales (ECACC).
La línea celular C6, procedente de células de glioma de rata, se adquirió de la Colección Europea de Cultivos Celulares Animales (ECACC).
En este trabajo, se emplearon sistemas de cocultivos entre las líneas celulares ECV304 y C6.Estas células crecen en medio M199 (Sigma) suplementado con Glutamax (Sigma-Aldrich), 10% de suero fetal de ternero (Gibco, Invitrogen Corporation) y penicilina/estreptomicina (100 U/mL; 100 μg/mL) (Gibco, Invitrogen Corporation). Las células se mantienen en atmósfera húmeda de 5% CO2/95% aire a 37ºC.
2.3. Métodos para el estudio de estrés oxidativo y disfunción mitocondrial
En el presente resumen de Tesis Doctoral se muestra una pequeña parte de los métodos empleados en el estudio de estrés oxidativo y disfunción mitocondrial.
- Medida de la generación intracelular de radicales libres: Técnica de diclorofluoresceína (DCFH-DA) (19). Tras los tratamientos, se añaden 200 μL/pocillo de una mezcla de una solución de 0,01 mM DCFH-DA con PBS glucosado (180 mg glucosa/100 mL PBS estéril) y se mantiene la placa en incubación durante 30 minutos a 37ºC. Después de este tiempo, las células se lavan con 200 μL/pocillo de PBS glucosado y se procede a la lectura utilizando un Fluorímetro Lector de microplacas FLx800, BioTek, Instruments, Inc., en las condiciones de lectura de excitación 480 nm y de emisión 510 nm.
- Determinación de la peroxidación lipídica (20). Para la cuantificación de la peroxidación lipídica se miden los niveles de MDA presentes en cada muestra mediante cromatografía líquida de alta eficacia (HPLC), siguiendo un protocolo modificado de Grotto et al. (2007).
- Determinación de la actividad y expresión de enzimas antioxidantes. Las enzimas estudiadas han sido aquellas implicadas en los principales procesos de detoxificación de ROS: catalasa (21), superóxido dismutasa (22), glutatión reductasa (23) y glutatión peroxidasa (24). Se ha determinado tanto la actividad por técnicas espectofotométricas así como la expresión proteica de cada una de ellas mediante Western blot.
- Determinación de los niveles de calcio mitocondrial. La concentración de calcio mitocondrial se determina midiendo la fluorescencia de Rhod-2.Después de los tratamientos, las células se lavan con medio de Krebs con calcio y sin sonda fluorescente, y a continuación se incuban durante 40 minutos a 37ºC en oscuridad con medio de Krebs con calcio y con la sonda fluorescente. Después de la incubación, las células se lavan 2 veces con medio de Krebs sin calcio y sin sonda fluorescente y se dejan incubar durante 30 minutos a la misma temperatura para permitir la hidrólisis completa del éster. La fluorescencia se mide a 37ºC durante 5 minutos a una λexcitación de 552 nm y a una λemisión de 581 nm a 37ºC en un fluorímetro Spectramax Gemini EM (Molecular Devices). A continuación, se adiciona el ionóforo A23187 (5 μM) y se mide la fluorescencia durante 15 minutos en las mismas condiciones descritas anteriormente (25).
2.4. Métodos para el estudio de paso a través de BHE
- Estudios de permeabilidad. El experimento de transporte se inicia con la adición de los diterpenos en un volumen de 0,5 mL en el compartimento apical (para el transporte apical a basal) y en un volumen de 1,5 mL en el compartimento basal (para el transporte basal a apical). Las placas se incuban a 37ºC en agitación durante 6 h. Una vez finalizado el transporte, se toman alícuotas de los lados apical y basal, para su posterior análisis y cuantificación por HPLC (26).
3. resultados y discusión
En el presente trabajo de Tesis, se ha evaluado la actividad neuroprotectora de seis diterpenos andalusol, conchitriol, lagascatriol, foliol, linearol y sidol aislados del género Sideritis frente al estrés oxidativo inducido por exposición a H2O2 en los modelos celulares PC12 (modelo neuronal) y U373-MG (modelo de astrocitoma).
Para llevarlo a cabo hemos establecido, en primer lugar, las condiciones de daño oxidativo para pasar posteriormente a determinar si las moléculas en estudio protegen frente al mismo. En el estudio sobre protección celular se ha evaluado el efecto sobre la viabilidad celular, los cambios sobre la morfología celular, el nivel de ROS intracelular, los cambios sobre los marcadores de estrés oxidativo (peroxidación lipídica y ratio GSH/GSSG) y sobre la actividad y expresión de las proteínas de las principales enzimas antioxidantes implicadas en el sistema de defensa. El efecto de los diterpenos sobre diferentes marcadores de disfunción mitocondrial como son los niveles de calcio citosólico y mitocondrial, el potencial de membrana mitocondrial y los niveles de ATP también han sido evaluados. Los posibles mecanismos implicados en la protección de los diterpenos en estudio frente al daño celular inducido por H2O2 han sido asimismo elucidados. Finalmente, se ha realizado el estudio y posible mecanismo de paso de los diterpenos a través de la BHE.
En el presente resumen de Tesis Doctoral se muestra sólo una pequeña parte de los resultados obtenidos.
El H2O2 ha sido la molécula utilizada como agente generador de estrés oxidativo. La formación de elevadas concentraciones de H2O2 se ha descrito en las fibrillas de amiloide formadas por la proteína beta-amiloide en la enfermedad de Alzheimer y, por la proteína alfa-sinucleína en la enfermedad de Parkinson, implicándose a esta especie reactiva de oxígeno como posible mecanismo causal en la muerte celular que ocurre en ambas enfermedades neurodegenerativas (27, 28).Teniendo en cuenta que el principal mecanismo de daño celular inducido por H2O2 implica la vía del estrés oxidativo (29), procedimos a estudiar el efecto de la exposición a H2O2 sobre el nivel de ROS intracelular mediante el ensayo de la diclorofluoresceína.
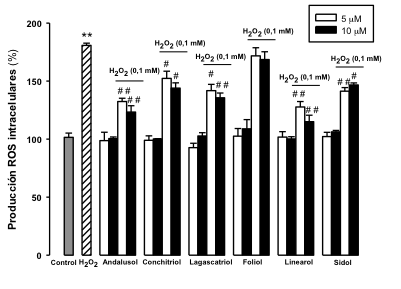
Como se muestra en la Figura 1, los niveles intracelulares de ROS se ven aumentados de manera significativa en comparación con las células control, lo que confirma que el H2O2, en las condiciones experimentales establecidas, es un inductor de estrés oxidativo. El pretratamiento de las células PC12 y U373-MG con los diterpenos ocasiona una inhibición de la producción de ROS intracelular, de manera significativa para andalusol, lagascatriol, linearol y sidol en los astrocitos y además para conchitriol, en las neuronas a las concentraciones de 5 y 10 μM.
A)
B)
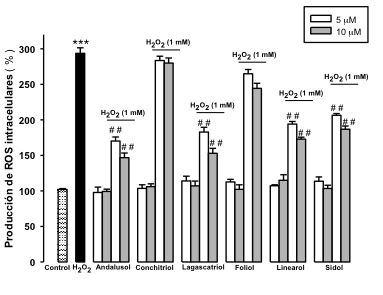
Figura 1.- Efecto sobre la producción de ROS intracelular. A) Células PC12. B) Células U373-MG. Los resultados se expresan como % de producción de ROS intracelulares respecto al control. Los resultados se expresan como media ± Desviación Estándar (n=9) de tres experimentos independientes. ** p < 0,01 versus control; # p < 0,05 y ## p < 0,01 versus H2O2
Una vez demostrada la capacidad de inducción de ROS por H2O2, se ha evaluado el efecto protector sobre los lípidos, componentes fundamentales de las membranas y una, y quizás la primera, diana biológica de los radicales libres. El daño en la membrana se ha determinado midiendo los niveles del producto final de la peroxidación lipídica malonildialdehido (MDA), ampliamente utilizado como biomarcador de estrés oxidativo en varias enfermedades neurodegenerativas (30, 31). Los resultados de este estudio (Figura 2) indican que la exposición a H2O2 en células PC12 y U373-MG produce una peroxidación lipídica significativa comparado con las células control. El pretratamiento celular con los diterpenos objeto de estudio, en concreto andalusol y linearol (en células PC12) y, andalusol y lagascatriol (en células U373-MG) protege frente a la peroxidación de lípidos de membrana inducida por H2O2, lo que por un lado confirma que estos compuestos son capaces de inhibir el daño oxidativo inducido por especies reactivas como ROS y por otro lado, se demuestra la capacidad de estos compuestos para interaccionar y penetrar las bicapas lipídicas, lo que le confiere valor añadido a su actividad protectora.

Figura 2.- Efecto sobre la peroxidación lipídica. El grado de peroxidación lipídica se cuantifica midiendo los niveles de MDA en los extractos celulares totales por HPLC. Los resultados se expresan como nmoles/mg proteína. Los resultados se expresan como media ± Desviación Estándar de tres experimentos independientes. * p < 0,05 y ** p< 0,01 versus control; #p< 0,05 y ##p< 0,01 versus H2O2.
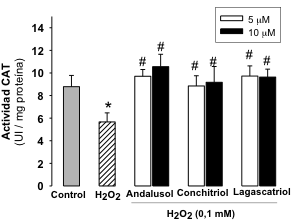
La activación de las enzimas antioxidantes es otro de los mecanismos que explica la acción de los compuestos antioxidantes. Cambios en los patrones enzimáticos de catalasa (CAT), superóxido dismutasa (SOD), glutation peroxidada (GPx), glutation reductasa (GR) y hemooxigenasa-1 (HO-1), entre otras, se traducen en variaciones del balance redox celular, constituyendo, por tanto, un biomarcador esencial del estrés oxidativo (32, 33). Así, con objeto de profundizar en el/los mecanismos de acción de los diterpenos del presente trabajo, procedimos a estudiar el papel de estos compuestos en la actividad y expresión de las enzimas antioxidantes, mediante técnicas espectrofotométricas y de Western-blot, respectivamente. En nuestro trabajo, el tratamiento de las células con H2O2 produce una significativa disminución tanto en la actividad como en la expresión de las enzimas anteriormente mencionadas. El pretratamiento celular con los diterpenos de Sideritis produce un aumento de la actividad y de la expresión de las enzimas antioxidantes endógenas evaluadas (excepto foliol en células PC12 y conchitriol y foliol en células U373-MG), comparado con las células expuestas a H2O2 como único tratamiento. En la Figura 3, se muestra como ejemplo la actividad y la expresión para la enzima catalasa y los compuestos andalusol, conchitriol y lagascatriol.
Modelos in vitro e in vivo han demostrado de forma evidente una asociación entre estrés oxidativo y disfunción mitocondrial. El aumento de los niveles de calcio mitocondrial puede llevar a un desacoplamiento de la función de la fosforilación oxidativa, y la consecuente disfunción mitocondrial. En la Figura 4, se muestra que ninguno de los diterpenos, a las concentraciones y tiempo de tratamiento ensayados, aumentan la concentración de calcio mitocondrial. Por otro lado, el H2O2 produce una significativa acumulación de calcio en la mitocondria. Cuando las células se tratan durante 24 h con los diterpenos objeto de estudio, previo a la exposición a H2O2, se observa una disminución en los niveles de calcio mitocondrial; concretamente, este efecto es significativamente marcado para linearol, andalusol (5 y 10 µM) y conchitriol (10 µM) en células PC12 y andalusol, lagascatriol y linearol (5 y 10 µM) y sidol (5 µM) en células U373-MG.
Una vez demostrado el potencial neuroprotector de los diterpenos en estudio, procedimos a evaluar si estos compuestos son capaces de atravesar la BHE. El estudio de permeabilidad (Tabla 1) revela que los valores de permeabilidad aparente de todos los diterpenos son menores que los del compuesto de referencia positivo, propranolol, y mayores que los del compuesto de referencia negativo, atenolol. Estudios previos han establecido que valores de permeabilidad (Papp) mayores de 3 x 10-6 cm/s indican que los compuestos pueden penetrar eficientemente la BHE (34). Por tanto, el valor de Papp de los diferentes diterpenos sugiere que éstos pueden atravesar la BHE vía transcelular por mecanismos de difusión pasiva. Andalusol y lagascatriol presentan los valores más altos de Papp, linearol y sidol tienen valores moderados y foliol y conchitriol, los más bajos.
A)
10 |
(mM) |
5 |
Control |
H2O2 |
5 |
Andalusol |
Conchitriol |
Lagascatriol |
H2O2 |
5 |
10 |
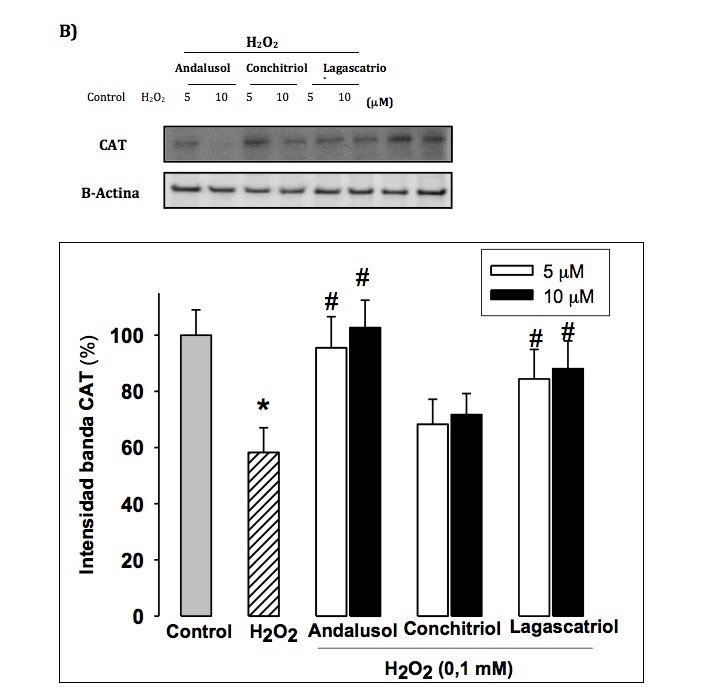
Figura 3.- Efecto en la actividad y expresión de la enzima CAT en células PC12. A) La actividad de la enzima CAT se cuantifica por métodos espectrofotométricos en extractos celulares totales. Los resultados de la actividad enzimática se expresan como UI/mg proteína. B) La expresión de proteínas se determina mediante Western blot en extractos celulares totales. Los resultados de la expresión de proteínas se expresan como % de intensidad de la banda, considerándose como 100% las células control. La ß-actina se utiliza como normalizador de carga. Se muestra una membrana representativa de la expresión de CAT (n=3). Los resultados se expresan como media ± Desviación Estándar de tres experimentos independientes. * p < 0,05 y ** p< 0,01 versus control; #p< 0,05 y ##p< 0,01 versus H2O2.
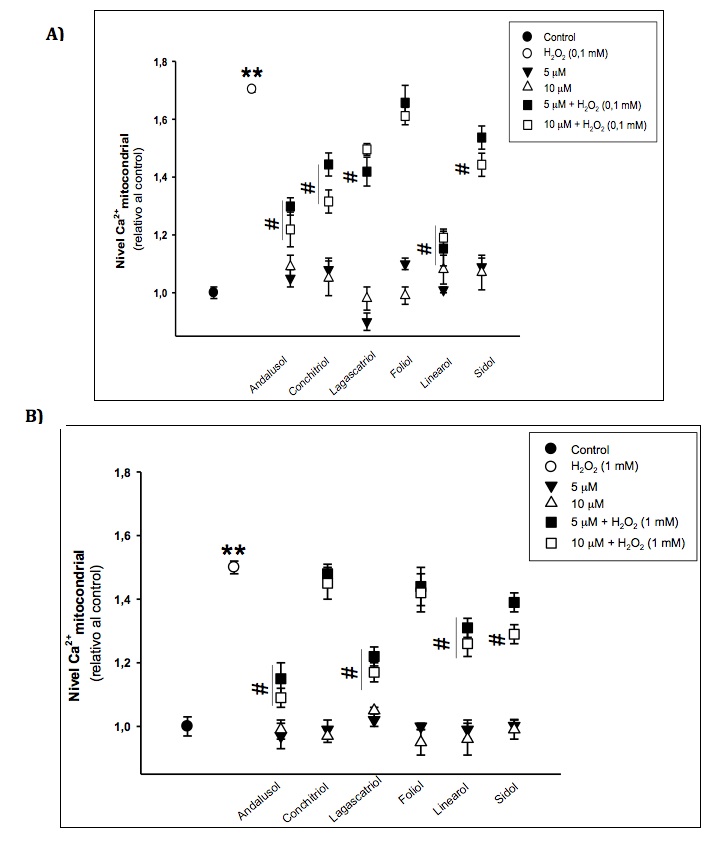
Figura 4.- Efecto sobre los niveles de calcio mitocondrial. Los cambios en los niveles de calcio mitocondrial se determinan por fluorescencia empleando el catión lipofílico fluorescente Rhod-2. Los resultados se expresan como media ± Desviación Estándar (n=9) de tres experimentos independientes. * p < 0,05 y ** p< 0,01 versus control; #p< 0,05 y ##p< 0,01 versus H2O2.
Tabla 1.- Estudios de permeabilidad. El transporte de los diferentes diterpenos se ha evaluado durante 6 horas en las direcciones absortiva(A-B) y secretora (B-A). Los compuestos de referencia propranolol y atenolol se utilizan como control positivo y negativo, respectivamente. El ratio se determina como Papp(B-A)/Papp(A-B). El porcentaje de recuperación se calcula como 100 – Mcélulas, siendo Mcélulas la cantidad de compuesto retenido en las membranas. ) [Mcélulas = MD0 – (MDt + ΣMR)] (ver material y métodos 2.3.2.4.). Los datos se expresan como media ± Desviación Estándar.
Compuesto |
Papp(x 10-6 cm/s) |
Pappcorregido(x 10-6 cm/s) |
Ratio |
Ratiocorregido |
% Recuperación |
||
A-B |
B-A |
A-B |
B-A |
||||
Andalusol |
9,1 ± 1,1 |
9,5 ± 2,8 |
11,4 ± 2,1 |
11,6 ± 2,6 |
0,9 |
1,0 |
78,8 ± 3,1 |
Conchitriol |
4,6 ± 0,9 |
4,8 ± 1,4 |
4,8 ± 1,1 |
4,9 ± 1,6 |
0,9 |
1,0 |
88,1 ± 2,2 |
Foliol |
3,8 ± 1,3 |
3,7 ± 1,9 |
4,0 ± 1,2 |
4,2 ± 1,5 |
0,9 |
0,9 |
90,5 ± 2,3 |
Lagascatriol |
8,8 ± 0,3 |
8,9 ± 0,5 |
10,7 ± 1,8 |
10,8 ± 1,5 |
1,0 |
1,0 |
81,5 ± 2,9 |
Linearol |
6,3 ± 0,5 |
6,6 ± 1,1 |
8,2 ± 1,1 |
8,3 ± 0,9 |
0,9 |
1,0 |
83,6 ± 3,3 |
Sidol |
6,9 ± 1,4 |
7, 2± 0,5 |
8,5 ± 1,7 |
8,6 ± 0,9 |
0,9 |
1,0 |
84,5 ± 2,8 |
Propranolol |
14,3 ± 1,1 |
14,8 ± 0,7 |
15,1 ± 1,3 |
15,6 ± 1,5 |
0,9 |
1,0 |
95,6 ± 1,9 |
Atenolol |
0,7 ± 0,4 |
0,8 ± 0,3 |
0,7 ± 0,6 |
0,8 ± 0,5 |
0,9 |
0,9 |
98,4 ± 2,1 |
4. Conclusiones
Los diterpenos estudiados ejercen un efecto neuroprotector frente al estrés oxidativo inducido por peróxido de hidrógeno, inhibiendo la producción de ROS, aumentando la actividad y la expresión de enzimas antioxidantes y atenuando los cambios derivados del estrés oxidativo a nivel mitocondrial.
De forma global, al comparar la eficiencia protectora de los 6 diterpenos de Sideritis spp. frente al estrés oxidativo inducido por H2O2, se observa un patrón de actividad similar pero no idéntico para todos estos compuestos en las dos líneas celulares estudiadas. Así, en la línea celular PC12, los diterpenos andalusol y linearol han sido los más activos, seguidos de sidol, lagascatriol y conchitriol, y por último, el compuesto menos activo ha sido foliol. En los estudios realizados sobre la línea celular U373-MG, los diterpenos que han ejercido el mayor efecto citoprotector han sido andalusol y lagascatriol, seguidos de linearol y sidol, y por último, de conchitriol y foliol.
5. agradecimientos
Este trabajo ha sido realizado en el Departamento de Farmacología de la Facultad de Farmacia de la Universidad Complutense de Madrid gracias a una beca predoctoral de Formación de Profesorado Universitario (FPU) del Ministerio de Ciencia e Innovación.
6. referencias
1. Ehrnhoefer, D.E.; Wong, B.K.; Hayden, M.R. Convergent pathogenic pathways in Alzheimer's and Huntington's diseases: shared targets for drug development. NatureReviewsDrug Discovery10, 853-867 (2011).
2. Katsuno, M., Tanaka, F., Sobue, G. Perspectives on molecular targeted therapies and clinical trials for neurodegenerative diseases. Journal of Neurology, Neurosurgery, and Psychiatry83, 329-335 (2012).
3. Barnham, K.J., Masters, C.L., Bush A.I. Neurodegenerative diseases and oxidative stress Nature Reviews Drug Discovery3, 205–214 (2004).
4. Hayashi, M., Miyata, R., Tanuma, N. Oxidative stress in developmental brain disorders.Advances in Experimental Medicine and Biology724, 278-290 (2012).
5. Valko, M., Morris, H., Cronin, M.T. Metals, toxicity and oxidative stress. Current Medicinal Chemistry12, 1161–1208 (2005).
6. Avery, S.V. Molecular targets of oxidative stress. Biochemical Journal434, 201–210 (2011).
7. Floyd, R.A. Antioxidants, oxidative stress, and degenerative neurological disorders. Proceedings of the Society for Experimental Biology and Medicine222, 236–245 (1999).
8. Ndhlala, A.R., Moyo, M., Van Staden, J. Natural antioxidants: fascinating or mythical biomolecules? Molecules 15, 6905-6930 (2010).
9. González-Burgos, E., Gómez-Serranillos, M.P. Terpene compounds in nature: A review of their potential antioxidant activity. Current Medicinal Chemistry19, 5319-5341 (2012).
10. Loğoğlu, E., Arslan, S., Öktemer, A., Şakiyan, İ. Biological activities of some natural compounds from SideritissipyleaBoiss. Phytotherapy Research20, 294-297 (2006).
11. González-Burgos, E., Carretero, M.E., Gómez-Serranillos, M.P. Sideritis spp.: Uses, chemical composition and pharmacological activities- A review. Journal ofEthnopharmacology135, 209-225 (2011).
12. Piozzi, F., Bruno, M., Rosselli, S., Maggio, A. The diterpenoids from the genus Sideritis. En: Atta-ur-Rahman (Ed) Studies in natural products chemistry 33, 493-540 (2006). Bioactive Natural Products. Elservier.
13. Venturella, P., Bellino, A. Constituents of Stachysitalica. Atti de ll’AccademiaNazionale di ScienzeArti Palermo IV 24, 95-99 (1965).
14. Piozzi, F., Venturella, P., Bellino, A., Mondelli, R. DiterpenesfromSideritissiculaUcria. Tetrahedron24, 4073-4081 (1968).
15. vonCarstenn-Lichterfelde, C., Valverde, S., Rodriguez, B. Studies on diterpenes from genus Sideritis. XIII two new stachene derivatives from Sideritisangustifolia (Labiatae). AustralianJournal of Chemistry27, 517–529 (1974).
16. Martín Panizo, F., Rodriguez, B., Valverde, S. Lagascatriol, a new diterpenoidfromSideritis angustifoliaLag. (Labiateae). Anales de Química 68, 1463-1465 (1972).
17. Martín Panizo, F., Rodríguez, B., Valverde, S. Estructura total revisada del lagascatriol, nuevo diterpeno de “Sideritis angustifolia” Lag. Anales de Química 70, 164-171 (1974).
18. García de Quesada, T., Rodríguez, B., Valverde, S. Six new diterpenesfromSideritisleucanthaCav. and Sideritislinearifolia Lam. TetrahedronLetter22, 2187-2190 (1972).
19. LeBel, C.P., Ischiropoulos H., Bondy S.C. Evaluation of the probe 2',7'-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chemical Research in Toxicology5, 227-231 (1992).
20. Grotto, D., Santa Maria, L.D., Boeira, S., Valentini, J., Charão, M.F., Moro, A.M., Nascimiento, P.C., Pomblum, V.J., Garcia, S.C. Rapid quantification of malondialdehyde in plasma by high performance liquid chromatography-visible detetion. Journal of Pharmaceutical and Biomedical Analysis 43, 619-624 (2007).
21. Aebi, H. Catalasein vitro. Methods in Enzymology105, 121-125 (1984).
22. Marklund, S., Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. European Journal of Biochemistry47, 469-474 (1974).
23. Barja de Quiroga, G., Perez-Campo, R. López-Torres, M. Anti-oxidant defences and peroxidation in liver and brain of aged rats. Biochemical Journal 272, 247-250 (1990).
24. Paglia, D.E., Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. Journal of Laboratory and Clinical Medicine 70, 158- 169 (1967).
25. Deniaud, A., Sharaf el dein, O., Maillier, E., Poncet, D., Kroemer, G., Lemaire, C., Brenner, C. Endoplasmic reticulum stress induces calcium-dependent permeability transition, mitochondrial outer membrane permeabilization and apoptosis. Oncogene 27, 285–299 (2008).
26. Gómez-Serranillos, P., Carretero, E., Slowing, K., Palomino, O. M., Villarrubia, A. I., Villar, A. HPLC Quantitative analysis of diterpenoids in Sideritis (Labiatae) species. Phytotherapy Research. 72, S101-S103 (1998).
27. Tabner, B.J., Turnbull, S., El-Agnaf, O., Allsop, D. Production of reactive oxygen species from aggregating proteins implicated in Alzheimer's disease, Parkinson's disease and other neurodegenerative diseases. Current Topics in Medicinal Chemistry1, 507-517 (2001).
28. Tabner, B.J., Turnbull, S., El-Agnaf, O.M., Allsop, D. Formation of hydrogen peroxide and hydroxyl radicals from A(beta) and alpha-synuclein as a possible mechanism of cell death in Alzheimer's disease and Parkinson's disease. Free Radical Biology and Medicine. 32, 1076-1083 (2002).
29. Halliwell, B. Reactive oxygen species and the central nervous system. Journal of Neurochemistry 59, 1609-1623 (1992).
30. Reed, T.T. Lipid peroxidation and neurodegenerative disease. Free Radical Biology and Medicine 51, 1302-1319 (2011).
31. Hayashi, M., Miyata, R., Tanuma, N. Oxidative stress in developmental brain disorders. Advances in Experimental Medicine and Biology 724, 278-290 (2012).
32. Wang, J.Y., Wen, L.L., Huang, Y.N., Chen, Y.T., Ku, M.C. Dual effects of antioxidants in neurodegeneration: direct neuroprotection against oxidative stress and indirect protection via suppression of glia-mediated inflammation. Current Pharmaceutical Design 12, 3521-333 (2006).
33. Jazwa, A., Cuadrado, A. Targeting heme oxygenase-1 for neuroprotection and neuroinflammation in neurodegenerative diseases. Current Drug Targets 11, 1517-1531 (2010).
34. Madgula, V.L., Avula, B., Yu, Y.B., Wang, Y.H., Tchantchou, F., Fisher, S., Luo, Y., Khan, I.A., Khan, S.I. Intestinal and blood-brain barrier permeability of ginkgolides and bilobalide: in vitro and in vivo approaches. PlantaMedica76, 599-606 (2010).