REVISIÓN |
Tratamientos experimentales contra el virus Ébola Zaire
Mª del Carmen Avendaño López1
1Real Academia Nacional de Farmacia, Académica de Número
e-mail: avendano@ucm.es
An. Real Acad. Farm. Vol. 80, Nº 4 (2014), pag.649-665
resumen
Se analizan las características de los filovirus, en especial del virus Ebola Zaire, así como varios tratamientos experimentados hasta el momento. Se incluyen vacunas, anticuerpos monoclonales, proteína C activada, proteína c2 recombinante de nematodos, interferones, RNAs de interferencia, oligonucleótidos antisentido (fósosforodiamidato morfolino oligómeros), y antimetabolitos (inhibidores de RNA polimerasa RNA-dependiente). Hacemos hincapié en los fundamentos, dianas erapéuticas, mecanismos de acción, objetivos y situación actual. También se comentan algunos antivirales descubiertos por métodos de cribado al azar y proyectos para un futuro inmediato. |
Palabras clave: Ébola; vacunas; tratamientos experimentales.
abstract
Experimental treatments against Zaire Ebola virus
The characteristics of filoviruses of the genus Ebolavirus, especially of Zaire Ebola virus, as well as several experimental treatments are analyzed. We include vaccines, monoclonal antibodies, recombinant human activated protein C, recombinant nematode anticoagulant protein c2, interferons, RNA interference, antisense ologonucleotides (phosphorodiamidate morpholino oligomers), and antimetabolites (RNA polimerase RNA-dependent inhibitors). We emphasize the fundamental principles, drug targets, action mechanisms, aims, and actual situation. We also comment some antiviral compounds found by throughput screening and near projects. |
Keywords: Ebola virus; vaccines; experimental treatments.
1. INTRODUCcióN
El jueves 30 de octubre de 2014, la Real Academia Nacional de Farmacia celebró una tertulia científica con el tema: "EBOLA: breve recapitulación sobre etiología, epidemiología, patogenia, tratamiento y control", en la que actuó como ponente el Prof. Dr. Rafael Rotger Anglada, Catedrático de Microbiología y Director del Centro de Análisis Sanitarios de la UCM. En su documentada intervención mencionó el tratamiento que según fuentes sanitarias se administró a la auxiliar de clínica Teresa Romero, la primera europea contagiada fuera de África que, afortunadamente, fue felizmente curada. Este acto, así como la avalancha de información producida como consecuencia del actual brote de Ébola (1), nos ha impulsado a elaborar una revisión que contemple gran parte de los esfuerzos realizados para la prevención y el tratamiento de esta epidemia.
2. CARACTERÍSTICAS DEL VIRUS ÉBOLA ZAIRE
Dentro de los filovirus, denominados así por su aspecto filamentoso, el género Ebolavirus comprende cinco serotipos o especies. A excepción de la especie Ebola Reston encontrada en Filipinas, todas las demás producen fiebre hemorrágica y una alta tasa de mortalidad en las personas. La especie Ebola Zaire (EBO-Z) tiene un periodo de incubación de 3-21 días, tras el cual los pacientes presentan síntomas gripales hasta que se produce una progresión muy rápida caracterizada por hemorragias y finalmente un fracaso multiorgánico (2). El primer brote documentado de esta especie se produjo en 1976 en la República Democrática del Congo, anteriormente denominada Zaire, y aunque desde entonces ha causado 34 epidemias en distintos países del África sub-Sahariana, ninguna ha causado la alarma de la actual por su posible diseminación al resto del mundo.
Un grave problema a la hora de estudiar vacunas o fármacos contra este virus es disponer de modelos animales adecuados ya que, aunque esta especie es letal para los ratones recién nacidos, los ratones adultos (que suelen ser los modelos más comunes para los estudios in vivo) son resistentes a la infección, por lo que si se utilizan para estos propósitos hay que manipularlos (3). Los macacos son por el momento el mejor modelo animal.
El genoma del virus Ébola consiste en una molécula única de ARN monocatenario lineal de polaridad negativa, por lo que no sirve directamente de ARN mensajero sino que mediante una RNA polimerasa vírica debe originar primero una copia complementaria que traduce las siete proteínas que posee el virión. Este está formado por un nucleoide con forma tubular rodeado por una cápsida helicoidal (Figura 1). El nucleoide está constituido por una nucleoproteína (N) y la mencionada RNA polimerasa (L). La cápsida está recubierta de una membrana en la que se integra una glicoproteína (GP), y en ella se encuentran también la proteína VP30 (que permite al virión desdoblarse dentro de una célula hospedadora), la VP35, y las proteínas VP24 y VP40 (que forman una matriz que mantiene unidos el nucleoide y la cápsida).
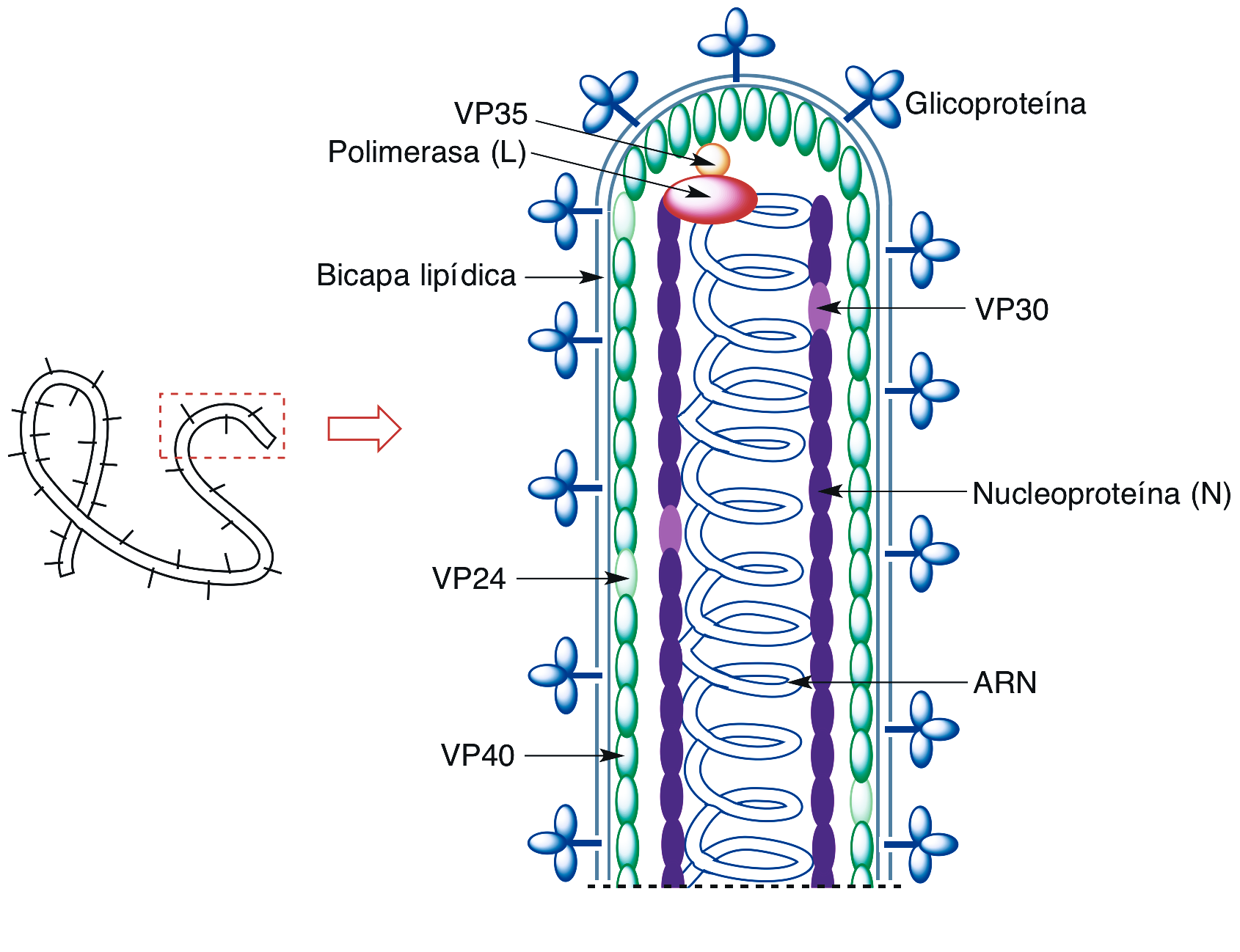
Figura 1.- Representación de un virión Ébola.
Los filovirus penetran en el citoplasma de la célula hospedadora a través de la fusión de su membrana externa con la membrana de endosomas, compartimentos de transporte desde la membrana plasmática a los lisosomas. En este proceso son especialmente importantes las glicoproteínas presentes en dicha membrana que, al experimentar procesos de proteolisis y de reordenamiento, facilitan dicha fusión y la posterior internalización del virus. El éxito obtenido en el desarrollo de antivirales y anticuerpos que tienen como objetivo impedir la entrada al interior celular de virus patógenos, incluyendo los virus de la gripe o el HIV, ha requerido un conocimiento detallado de los correspondientes mecanismos de entrada. Este conocimiento ha demostrado que muchas de las proteínas virales que intervienen en la fusión e internalización de distintos virus poseen lazos con residuos hidrófobos (fusion loops, FL). En el caso del virus Ébola Zaire se ha determinado la estructura de estos lazos (4) (Figura 2) y cuáles son los residuos de aminoácidos que son críticos (5).
Estudios in vitro también han demostrado que un receptor de la glicoproteína
vírica es la proteína Niemann-Pick C1 (NPC1), conocida por su importante papel
en el transporte de colesterol (6). En estos estudios se utilizaron como
inhibidores de NPC1 derivados de adamantano, por ejemplo el compuesto 1, observándose que cuanto menor es la
capacidad para transportar el colesterol mayor es la capacidad para resistir la
infección del virus del Ébola.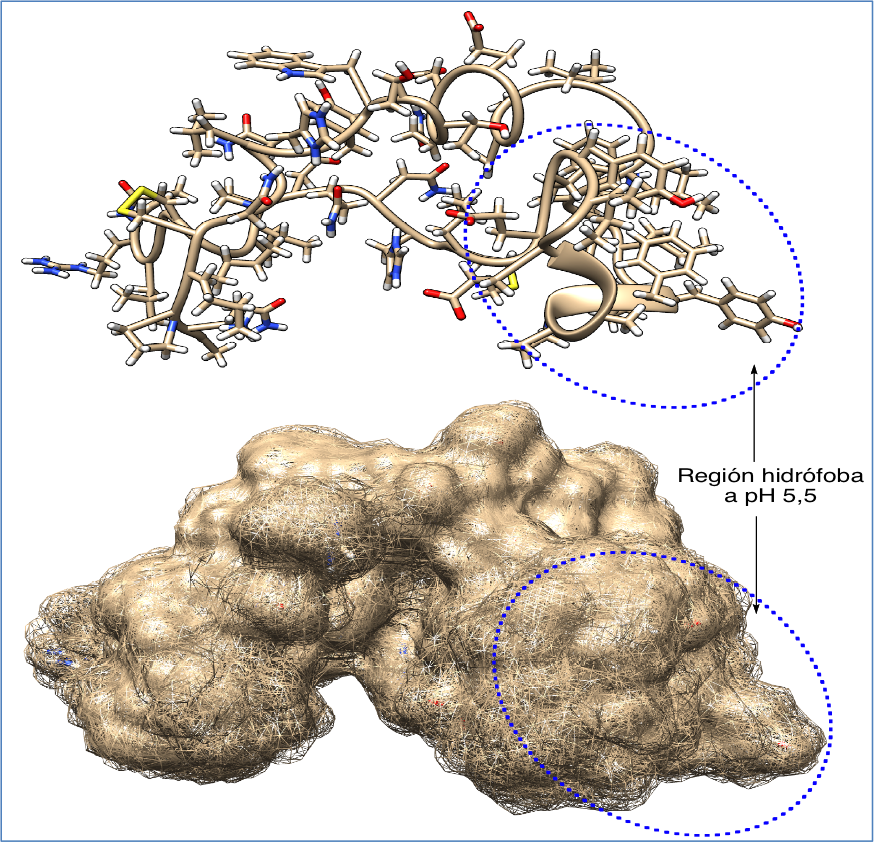
Figura 2.- Estructura de un lazo de fusión del virus Ébola, generada a partir de pdb 2LCY y representada con Chimera 1.81.
Sin embargo, este hallazgo no ha conducido por el momento a ninguna actuación terapéutica, ya que el uso de estos inhibidores induciría la enfermedad denominada Niemann-Pick de tipo C, una enfermedad rara de origen hereditario con una prevalencia de un caso por cada 150.000 habitantes que es consecuencia de mutaciones en las proteínas NPC1 y NPC2. Esta enfermedad afecta fundamentalmente a niños de edad escolar y conduce a un desenlace fatal por la acumulación de colesterol en los lisosomas de tejidos y órganos (7).
Tras la entrada en el citoplasma, el genoma vírico se replica y transcribe, se sintetizan las correspondientes proteínas y brotan nuevos viriones al exterior (Figura 3).
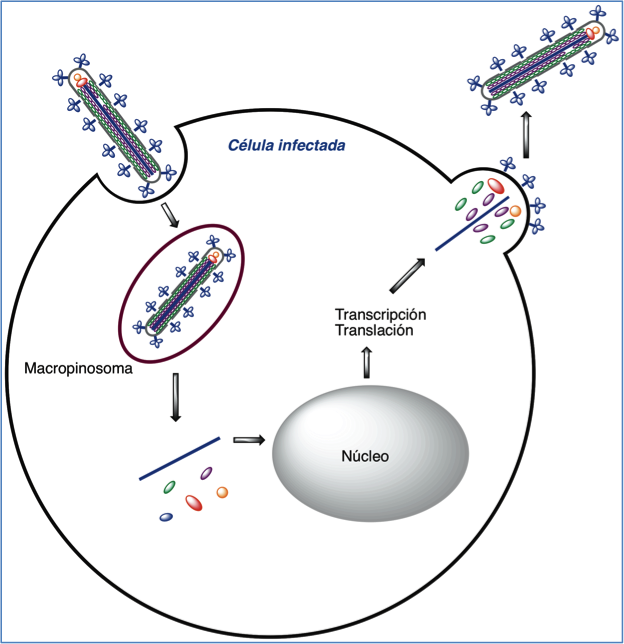
Figura 3.- Ciclo vital del virus Ébola.
3. tratamientos experimentales. vacunas
El desarrollo de vacunas eficaces es la mejor manera de contener y atajar la epidemia e incluso podría ser más fácil que para otros virus ARN como el VIH (8). Hasta el momento se han desarrollado múltiples vacunas experimentales que han demostrado su eficacia utilizando primates como animales modelo, pero no hay todavía ninguna aprobada para uso humano (Figura 4).
Figura 4.- Vacunas frente al virus Ébola Zaire |
Tampoco se ha aprobado hasta el momento ningún fármaco, ya sea molécula pequeña o biológica. Aunque se han investigado varios, muy pocos han iniciado ensayos clínicos en humanos. Por ello, el tratamiento de los enfermos en África se ha limitado al uso de cuidados paliativos. Estos cuidados, junto a los métodos empleados para prevenir la transmisión, todavía no han resuelto el actual brote. Comentaremos algunos de los tratamientos estudiados, centrándonos en los fundamentos en que se han basado.
4. ANTICUERPOS MONOCLONALES
El desarrollo de anticuerpos monoclonales (mAbs) anti-Ébola está basado en el mismo principio que las vacunas: la administración de anticuerpos que estimulen el sistema inmune del paciente para que responda de forma más rápida y eficaz al virus. Entre ellos se incluyen los mAbs quiméricos humanizados de ratón c13C6, h-13F6, y c6D8, cuya mezcla se denomina MB-300. Tras la administración de MB-300 a macacos una hora después de la infección por virus Ébola, estos animales mostraron una viremia baja o nula (10). Por otra parte, tres mAbs murinos: m1H3, m2G4 y m4G7, componentes de otro cóctel denominado ZMAb, se humanizaron para originar el cóctel de mAbs c1H3, c2G4 y c4G7 denominado cZMAb. Estos anticuerpos, producidos en la planta Nicotiana benthamiana, dieron lugar a la mezcla denominada ZMapp, que ataca a proteínas de la superficie del virus (Figura 5).
ZMapp se desarrolla por la empresa Mapp Biopharmaceuticals y parece ser la mezcla de anticuerpos más eficaz hasta el momento, ya que es capaz de rescatar al 100% de los macacos cuando el tratamiento se inicia 5 días después de la infección (11). Sin embargo, su proceso de producción es muy lento, ya que depende del crecimiento y aislamiento de una planta, por lo que se han obtenido y utilizado hasta ahora cantidades tan pequeñas que no se sabe si las personas que han sobrevivido al tratamiento lo han hecho gracias a ZMapp o a sus propias defensas.
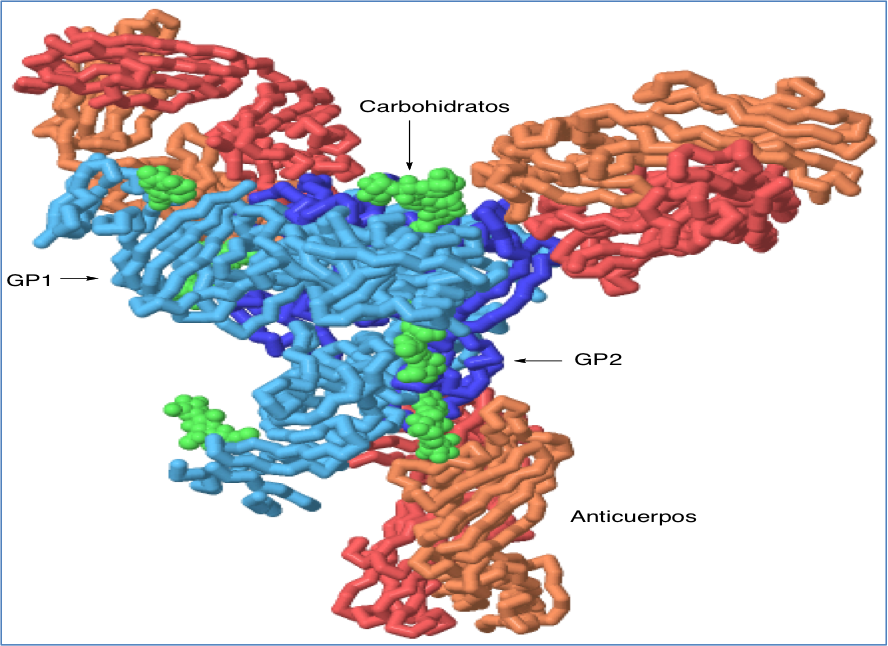
Figura 5.- Estructura del complejo que forma la glicoproteína del virus Ébola con anticuerpos humanos, aislado de un superviviente de la enfermedad. La estructura se ha generado a partir de pdb 3CSY y se ha representado con Jmol. Se observan dos cadenas en la glicoproteína, GP1 que participa en la unión a receptor y GP2 que se encarga de la fusión a la célula infectada. La estructura incluye algunas moléculas de carbohidratos, que en condiciones fisiológicas recubren completamente la proteína.
5. PROTEÍNA C ACTIVADA, PROTEÍNA C2 RECOMBINANTE DE NEMATODOS, E INTERFERONES
Además de las vacunas y de los cócteles de mAbs, diversas aproximaciones estudiadas en primates en la última década han demostrado ser de interés como posibles tratamientos del Ébola, aunque es su mayoría sólo producen efectos beneficiosos si se administran antes del segundo día post-infección. Estas aproximaciones incluyen la administración de la proteína C activada recombinante humana (rhAPC) o de la proteína c2 recombinante de nematodos (rNAPc2), y la administración de interferones.
La proteína C activada recombinante humana (rhAPC, Xigris®), fabricada por la empresa Eli Lilly, estaba autorizada cuando existía grave riesgo de muerte por sepsis severas (12). En estas circunstancias, hay una rápida disminución de los niveles de proteína C activada, al igual que ocurre en la fiebre hemorrágica causada por filovirus. Por esta razón, se suministró a macacos una infusión intravenosa de rhAPC a los 30-60 minutos del contagio y se continuó el tratamiento durante 7 días, demostrándose que la fiebre hemorrágica de origen vírico y las sepsis severas pueden beneficiarse de tratamientos comunes (13).
Por su parte, el estudio de la proteína c2 recombinante de nematodos (rNAPc2) pretendía averiguar si esta enzima, que es una proteasa de serina inhibidora del factor VII iniciador de la coagulación de la sangre, es capaz de evitar la coagulación intravascular diseminada que produce el virus. Su administración a macacos infectados tras 10 minutos o tras 24 horas de la infección prolongó el tiempo de supervivencia, lo que se interpretó como un punto de partida de los tratamientos dirigidos al contrarrestar los efectos de la enfermedad en vez de los dirigidos a inhibir la replicación vírica (14).
En cuanto al uso de interferones, hemos de recordar que los interferones a y b son citocinas que al interaccionar con su receptor (interferon-α/β receptor, IFNAR) activan la vía de señalización JAK/STAT y producen una respuesta inmunológica especialmente dirigida contra los virus (Figura 6) (15).
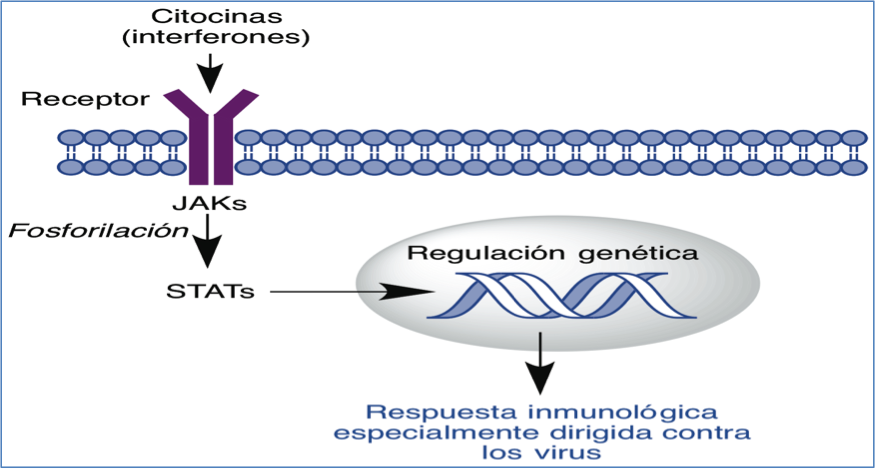
Figura 6.- Respuesta inmune inducida por la unión de los interferones a/b a su receptor transmitida por la vía de señalización JAK/STAT.
En la fiebre hemorrágica producida por filovirus se observan concentraciones de interferón b en plasma que son 60-100 veces superiores a las que existen en otras infecciones víricas, junto a una baja producción del mismo. Estos datos sugieren que la terapia con interferón podría utilizarse en el tratamiento de las infecciones causadas por estos virus y, en efecto, la administración de interferón b a macacos recientemente infectados prolongó su supervivencia (16). Por otra parte, la combinación de Zmab con interferón a (Ad-IFN) utilizando adenovirus como vectores del mismo, ha demostrado ser eficaz en primates si el tratamiento se inicia pasados 3 días desde la infección. Este aumento en la ventana terapéutica permitiría su uso en el momento en que la viremia y sus síntomas son detectables (17).
6. RNAs de INTERFERENCIA y OLIGONUCLEÓTIDOS ANTISENTIDO
Hay otros enfoques terapéuticos que están basados en la administración de RNAs de interferencia (siRNA) y oligonucleótidos antisentido (ASOs) de tipo PMOplus (positively-charged phosphorodiamidate morpholino oligomers).
El uso de RNAs de interferencia (siRNA) contra la RNA polimerasa (L) viral se ensayó primero en cobayas, a los que se administró una formulación estable de aquéllos inmediatamente después de la infección. Los resultados favorables indujeron a un posterior estudio en primates utilizando una combinación de siRNAs formulados de la misma manera que tenían como diana a las proteínas víricas polimerasa (L), VP24 y VP35. Un grupo de macacos, a los que se administró esta mezcla a los 30 minutos de la infección y en los 6 días siguientes, experimentó una protección del 66%. Estos datos se interpretaron como una demostración del potencial de los RNA de interferencia (18).
Por su parte, los oligonucleótidos antisentido tienen aquí como diana determinadas secuencias de RNA, copia del RNA viral, que son capaces de traducirse en proteínas. Por consiguiente, la inhibición de esta secuencias produce el bloqueo de este proceso (Figura 7).
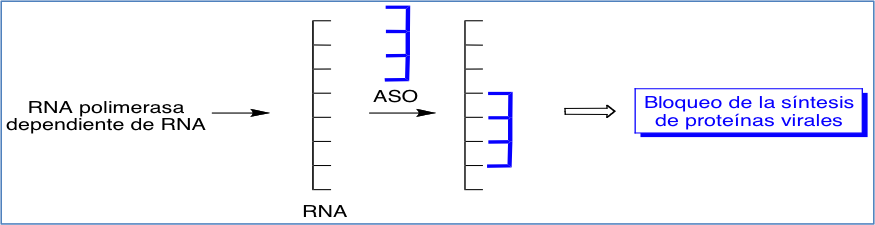
Figura 7. Bloqueo de la síntesis de proteínas virales producido por la acción de los nucleótidos antisentido (ASOs).
Dentro de esta aproximación hay que mencionar dos combinaciones de oligómeros de morfolino fósforodiamidato para al tratamiento de los virus Ébola y Marburg, respectivamente. Tras un estudio previo en primates, estas combinaciones han sido objeto de ensayos clínicos de fase I. AVI-6002 está dirigida a evitar la expresión de las proteínas de la matriz VP24 y VP35, mientras que AVI-6003 está dirigida a evitar la expresión de las proteínas VP24 y NP) (19). En ambos caso se ha utilizado la metodología química denominada PMOplus,® desarrollada por la firma Sarepta Therapeutics. Los componentes de AVI-6002 son AVI-7537 y AVI-7539, cada uno de los cuales es un oligómero de morfolino fósforodiamidato en el que ciertos agrupamientos de tipo dimetilamina se han sustituido por anillos de piperazina (ver la Figura 8). Recordemos que los oligonucleótidos con potencial terapéutico, además de tener una afinidad selectiva con la secuencia de RNA a la que se dirigen, tienen que evitar su degradación por las enzimas denominadas nucleasas. AVI-7537 y AVI-7539, debido a la presencia de residuos de piperazina, contienen cargas positivas que favorecen su interacción con el RNA vírico, que está negativamente cargado.

Figura 8.- Estructura general de algunos oligonucleótidos. Morfolino fósforodiamidatos componentes de AVI-7537 y AVI-7539.
7. ANTIMETABOLITOS INHIBIDORES DE RNA POLIMERASA RNA-DEPENDIENTE
El uso de antimetabolitos que inhiban la RNA polimerasa RNA-dependiente, o causen mutagénesis letales cuando se incorporen al RNA viral, es otro enfoque terapéutico de interés aunque, dada la facilidad de estos virus para sufrir mutaciones, estos fármacos podrían desarrollar resistencias y quizás requerirían su uso en combinación. Se conocen varios profármacos de estos antimetabolitos, como BCX4430, favipiravir y brincidofovir, que inhiben la polimerase de RNA dependiente de RNA de filovirus (Figura 9). El compuesto BCX4430 es un azanucleósido análogo de forodesina, un fármaco huérfano indicado para el tratamiento de varios tipos de leucemia, que se desarrolló primero como posible tratamiento de la hepatitis C y luego en infecciones por filovirus con la colaboración del NIAID. En estos estudios, BCX4430 demostró su actividad in vitro como terminador de cadena. Ensayos con roedores y macacos han demostrado que protege frente a virus Ébola y Marburg cuando se administra antes del tercer día tras la infección (20).

Figura 9.- Algunos profármacos de antimetabolitos que inhiben la polimerase de RNA dependiente de RNA de filovirus.
Favipiravir, también conocido como T-705, se descubrió en los laboratorios de investigación de la empresa Toyama Chemical Co., Ltd. Basándose en su actividad para inhibir la replicación del virus influenza en ratones, demostrada en estudios realizados en colaboración con el Profesor Kimiyasu Shiraki de la Universidad de Toyama, se seleccionó para ser estudiado clínicamente. También fue activo frente a diferentes tipos de virus ARN, entre otros el virus de la fiebre amarilla y el virus del Nilo Occidental. Su interés aumentó debido a que su novedoso mecanismo de acción (21) le permitió proteger a ratones de una infección letal de virus H5N1 altamente patógenos resistentes al inhibidor de neuraminidasa oseltamivir (22). Fabricado por el grupo Fuji Film, favipiravir se utiliza en Japón como antigripal con el nombre comercial Avigan,® y se encuentra en ensayos clínicos avanzados para ser aprobado para este indicación en EEUU (23). Las enzimas de la célula huésped lo bioactivan transformándolo en el ribofuranosil-5′-trifosfato correspondiente (T-705-RTP), que actúa probablemente como un falso nucleótido incorporándose al RNA viral e inhibiendo su RNA polimerasa dependiente de RNA (Figura 10). Se ha comprobado que antes de transformarse en el trifosfato, la enzima humana hipoxanthina guanina fosforibosiltransferasa (HGPRT) lo convierte en su ribosa-5′-monofosfato (T-705-RMP) (24).
Recientemente se ha empleado favipiravir experimentalmente para tratar la infección por el virus Ébola en humanos, basándose en estudios previos en los que mostró esta actividad in vitro e in vivo (25) (en los estudios in vivo se utilizaron ratones carentes del receptor de interferón tipo I, IFNAR-/-). Aunque se sabe que favipiravir no produce toxicidad aparente en las células de mamífero, ni inhibe en ellas la síntesis de DNA o RNA, se desconoce su seguridad y eficacia en las dosis empleadas contra el virus Ébola. Según fuentes médicas, en España se ha utilizado favipiravir en el tratamiento de Teresa Romero a dosis mucho mayores que las que se emplean para su uso como antigripal.

Figura 10.- Bioactivación de favipiravir y mecanismo de acción.
El liberiano diagnosticado en Dallas (Tejas) como enfermo de Ébola, se trató con brincidofovir (CMX001), un conjugado de cidofovir que se convierte intracelularmente en el antiviral activo cidofovir difosfato (cidofovir-DP). Gracias a su cadena lipídica, brincidofovir es un profármaco que puede administrase por vía oral, siendo más potente que cidofovir porque se encuentra a mayores concentraciones intracelulares y carece de la toxicidad renal de cidofovir. Este es un análogo de 2’desoxicitidina monofosfato originalmente desarrollado para el tratamiento de infecciones por virus DNA, que cuando penetra en el interior celular se disfosforila a cidofovir-DP y este, debido a su semejanza con citidina-TP, actúa como falso sustrato de polimerasas DNA inhibiendo finalmente la replicación (Figura 11) (26).

Figura 11.- Mecanismo de acción de brincidofovir.
Brincidofovir inhibe poxvirus, herpesvirus, y adenovirus, habiéndose estudiado en ensayos clínicos de fases II y III. Por su actividad frente a los poxvirus, se almacena en EEUU como defensa frente a un posible ataque bioterrorista con virus de la viruela. Cuando surgió la epidemia de Ébola en 2014, la firma Chimerix que lo fabrica lo puso (junto con otros de sus compuestos) a disposición de las autoridades sanitarias para averiguar si podía eliminar este virus, aunque no fuera un virus DNA. Inesperadamente, brincidofovir fue un potente inhibidor de Ébola in vitro, por lo que la FDA autorizó su uso con el paciente de Dallas que finalmente falleció.
8. ANTIVIRALES ENCONTRADOS POR CRIBADO AL AZAR
El cribado de moléculas pequeñas para observar en ellas una posible actividad antiviral ha permitido identificar varios inhibidores de filovirus. Por ejemplo, en un cribado in vitro de distintos fármacos conocidos, se encontró esta actividad en los moduladores del receptor estrogénico clomifeno y toremifeno (Figura 12).

Figura 12.- Inhibidores de filovirus hallados por métodos de cribado al azar.
Utilizando ratones modificados convenientemente modificados para poder ser utilizados como modelos de infección, estos fármacos fueron también activos in vivo, demostrándose que esta acción es independiente de los receptores de estrógenos y se debe a que interfieren el proceso de entrada del virus al interior celular anteriormente mencionado (27). El compuesto NSC 62914, que posee actividad in vitro e in vivo frente a los virus Ébola y Marburg entre otros, se encontró en el cribado de otras estructuras. Este compuesto es un antioxidante que atrapa especies reactivas de oxígeno, modulando así el estrés oxidativo. Sin embargo, dado que cuando se estudiaron otros compuestos antioxidantes no se observó ningún efecto antiviral, se cree que su acción antiviral debe implicar la modulación de múltiples dianas o vías de señalización (28).
9. PROYECTOS FUTUROS
“Médicos Sin Fronteras” ha anunciado que pretende realizar a partir de febrero de 2015 tres proyectos de investigación en sus centros africanos. Uno en Conakry, Guinea, liderado por el Antwerp Institute of Tropical Medicine (ITM) estudiará la administración de sangre y plasma de pacientes de Ébola que han sobrevivido y que, por tanto, contienen anticuerpos específicos. Otro estudiará, en un lugar todavía sin determinar, la administración dos veces por semana durante dos semanas de formulaciones orales de brincidofovir a 140 pacientes, y estará financiado por el Wellcome Trust y liderado por la Universidad de Oxford. El tercero, liderado por el French National Institute of Health and Medical Research (Inserm), se llevará a cabo en Gueckedou, Guinea, y estudiará el uso de favipiravir. En cuanto a las vacunas, aunque normalmente se requeriría más tiempo para su aprobación, se pretende que a principios del año 2015 lleguen a África y se utilicen unas 20.000 dosis de las vacuna producidas por GlaxoSmithKline (GSK) y la Agencia de Salud Pública de Canadá.
10. CONCLUSIONES
Aunque no hay todavía disponible para su uso humano ninguna vacuna o tratamiento para la fiebre hemorrágica causada por el virus Ébola, los conocimientos básicos y los hallazgos experimentales que ya existen permiten ser optimistas acerca de una aplicación clínica no muy lejana. Sin embargo, parece claro que no será posible este cambio de situación sin una importante aportación de fondos públicos a la financiación de este proceso.
10. CONCLUSIONS
Although neither a vaccine nor an effective antiviral treatment is currently available for use in humans, the fundamental knowledge and experimental findings already achieved, generate cautious optimism about the translation of these findings into clinical practice for prevention and treatment of Ebola hemorrhagic fever. However, it seems quite obvious that an effective progress will require public financial support.
11. Agradecimientos
La autora desea expresar su agradecimiento al Dr. J. C. Menéndez por la elaboración de algunas de las figuras.
12. referencias
1. Gatherer, D. The 2014 Ebola virus disease outbreak in West Africa. J. Gen. Virol. 95, 1619-1624 (2014).
2. Feldmann, H.; Geisbert, T. W. Ebola haemorrhagic fever. Lancet, 377, 849-862 (2011).
3. Ver, por ejemplo: (a) Bray, M.; Davis, K.; Geisbert, T.; Schmaljohn, C.; Huggins; J. A mouse model for evaluation of prophylaxis and therapy of Ebola hemorrhagic fever. J. Infect. Dis. 179, S248-58,(1999) (b) Bausch, D. G.; Sprecher, A. G.; Jeffs, B.; Boumandouki, P. Treatment of Marburg and Ebola hemorrhagic fevers: a strategy for testing new drugs and vaccines under outbreak conditions. Antiviral Res. 78, 150-161 (2008).
4. Gregory, S. M.; Harada, E.; Liang, B.; Delos, S. E.; White, J. M.; Tamm, L. K. Structure and function of the complete internal fusion loop from Ebolavirus glycoprotein 2. Proc. Natl. Acad. Sci. U S A. 108, 11211-11216 (2011).
5. Gregory, S. M.; Larsson, P.; Nelson, E. A.; Kasson, P. M.; White, J. M.; Tamm, L. K. Ebolavirus entry requires a compact hydrophobic fist at the tip of the fusion loop. J. Virol. 88, 6636-6649 (2014).
6. (a) Carette, J. E.; Raaben, M.; Wong, A. C.; Herbert, A. S.; Obernosterer, G.; Mulherkar, N.; Kuehne, A. I.; Kranzusch, P. J.; Griffin, A. M.; Ruthel, G.; Dal Cin, P.; Dye, J. M.; Whelan, S. P.; Chandran, K.; Brummelkamp, T. R. Ebola virus entry requires the cholesterol transporter Niemann–Pick C1. Nature 477, 340-343 (2011). (b) Côté, M.; Misasi, J.; Ren, T.; Bruchez, A.; Lee, K.; Filone, C. M.; Hensley, L.; Li, Q.; Ory, D.; Chandran, K.; Cunningham, J. Small molecule inhibitors reveal Niemann-Pick C1 is essential for Ebola virus infection. Nature, 477, 344-348 (2011). (c) White, J. M.; Schornberg, K. L. A new player in the puzzle of filovirus entry. Nat. Rev. Microbiol. 10, 317-322 (2012).
7. Chang, T.-Y.; Reid, P. C.; Sugii, S.; Ohgami, N.;Cruz, J. C.;. Chang, C. C. Y. Niemann-Pick Type C Disease and Intracellular Cholesterol Trafficking. J. Biol. Chem. 280, 20917-20920 (2005).
8. Dhillon, R. S.; Srikrishna, D.; Sachs, J. Controlling Ebola: next steps Lancet 384, 1409-1411 (2014).
9. Kanapathipillai, R.; Restrepo, A. N. H.; Fast, P.; Wood, D.; Dye, Ch.; Kieny, M.-P.; Moorthy, V. Ebola Vaccine-An Urgent International Priority. N. Eng. J. Med. DOI: 10.1056/NEJMp1412166 (2014).
10. Olinger, G. G.; Pettitt, J.; Kim, D.; Working, C.; Bohorov, O.; Bratcher, B.; Hiatt, E.; Hume, S. D.; Jhonson, A. K.; Morton, J.; Pauly, M.; Whaley, K. J.; Lear, C. M.; Biggins, J. E.; Scully, C.; Hensley, L.; Zeitlin, L. Delayed treatment of Ebola virus infection with plant-derived monoclonal antibodies provides protection in rhesus machaques. Proc. Natl. Acad. Sci. U S A. 109, 18030-18035 (2012).
11. Qiu, X.; Wong, G.; Audet, J.; Bello, A.; Fernando, L.; Alimonti, J. B.; Fausther-Bovendo, H.; Wei, H.; Aviles, J.; Hiatt, E.; Jhonson, A.; Morton, J.; Swope, K.; Bohorov, O.; Bohorova, N.; Goodman, C.; Kim, D.; Pauly, M. H.; Velasco, J.; Pettitt, J.; Olinger, G. G.; Whaley, K.; Xu, B.; Strong, J. E.; Zeitlin, L.; Kobinger, G. P. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 514, 47-53 (2014).
12. La empresa retiró este producto del mercado en octubre de 2011.
13. Hensley, L. E.; Stevens, E. L.; Yan, S. B.; Geisbert, J. B.;. Macias, W. L.; Larsen, T.; Daddario-DiCaprio, K. M.; Cassell, G. H.; Jahrling, P. B.; Geisbert, T. W. Recombinant human activated protein C for the postexposure treatment of Ebola hemorrhagic fever. J. Infect. Dis. 196, S390-S399 (2007).
14. Geisbert, T. W.; Hensley, L. E.; Jahrling, P. B.; Larsen, T.; Geisbert, J. B.; Paragas, J.; Young, H. A.; Fredeking, T. M.; Rote, W. E.; Vlasuk, G. P. Treatment of Ebola virus infection with a recombinant inhibitor of factor VIIa/tissue factor: a study in rhesus monkeys. Lancet 362, 1953-1958 (2003).
15. Uzé, G.; Schreiber, G.; Piehler, J.; Pellegrini, S. The receptor of the type I interferon family. Curr. Top. Microbiol. Immunol. 316, 71-95 (2007).
16. Smith, L. M.; Hensley, L. E.; Geisbert, T. W.; Johnson, J.; Stossel, A.; Honko, A.; Yen, J. Y.; Geisbert, J.; Paragas, J.; Fritz, E.; Olinger, G.; Young, H. A.; Rubins, K. H.; Karp, C. L. Interferon-beta therapy prolongs survival in rhesus macaque models of Ebola and Marburg hemorrhagic fever. J. Infect. Dis. 208, 310-318 (2013).
17. Qiu, X.; Wong, G.; Fernando, L.; Audet, J.; Bello, A.; Strong, J.; Alimonti, J. B.; Kobinger, J. P. MAbs and Ad-vectored IFN-alpha therapy rescue Ebola-infected nonhuman primates when administered after the detection of viremia and symptoms, Sci. Transl. Med. 5, 207 ra143 (2013).
18. Geisbert, T. W.; Lee, A. C.; Robbins, M.; Geisbert, J. B.; Honko, A. N.; Sood, V.; Johnson, J. C.; de Jong, S.; Tavakoli, I.; Judge, A.; Hensley, L. E.; Maclachlan, I. Postexposure protection of non-human primates against a lethal Ebola virus challenge with RNA interference: a proof-of-concept study. Lancet, 375, 1896-1905 (2010).
19. Heald, A. E.; Iversen, P. L.; Saoud, J. B.; Sazani, P.; Charleston, J. S.; Axtelle, T.; Wong, M.; Smith, W. B.; Vutikullird, A.; Kaye, E. Safety and Pharmacokinetic Profiles of Phosphorodiamidate Morpholino Oligomers with Activity against Ebola Virus and Marburg Virus: Results of Two Single-Ascending-Dose Studies. Antimicrob. Agents Chemother, 58, 6639-6647 (2014).
20. (a) Stock, I. Marburg and Ebola hemorrhagic fevers-pathogens, epidemiology and therapy. Med. Monatsschr. Pharm. 37, 324-330 (2014). (b) Warren, T. K.; Wells, J.; Panchal, R. G.; Stuthman, K. S.; Garza, N. L.; Van Tongeren, S. A.; Dong, L.; Retterer, C. J.; Eaton, B. P.; Pegoraro, G.; Honnold, S.; Bantia, S.; Kotian, P.; Chen, X.; Taubenheim, B. R.; Welch, L. S.; Minning, D. M.; Babu, Y. S.; Sheridan, W. P.; Bavari, S. Protection against filovirus diseases by a novel broad-spectrum nucleoside analogue BCX4430. Nature 508, 402-405, (2014).
21. Baranovich, T.; , S.-S.; Armstrong, J.; Marjuki, H.; , R. J.; Webster, R. G.; Govorkova, E. A. T-705 (Favipiravir) Induces Lethal Mutagenesis in Influenza A H1N1 Viruses in Vitro. J. Virol. 87, 3741-3751 (2013).
22. Kiso, M.; Takahashi, K.; Sakai-Tagawa, Y.; Shinya, K.; Sakabe, S.; Le, Q. M.; Ozawa, M.; Furuta, Y.; Kawaoka, Y. T-705 (favipiravir) activity against lethal H5N1 influenza A viruses. Proc. Natl. Acad. Sci. U S A 107, 882-887 (2010).
23. (a) Furuta, Y.; Takahashi, K.; Fukuda, Y.; Kuno, M.; Kamiyama, T.; Kozaki, K.; Nomura, N.; Egawa, H.; Minami, S.; Watanabe, Y.; Narita, H.; Shiraki, K. In vitro and in vivo activities of anti-influenza virus compound T-705. Antimicrob. Agents Chemother. 46, 977-981 (2002). (b) Para una revisión ver: Furuta, Y.; Gowen, B. B.; Takahashi, K.; Shiraki, K.; Smee, D. F.; Barnard, D. L. Favipiravir (T-705), a novel viral RNA polymerase inhibitor. Antiviral Res. 100, 446-454 (2013).
24. Naesens, L.; Guddat, L. W.; Keough, D. T.; van Kuilenburg, A. B.; Meijer, J.; Vande Voorde, J.; Balzarini, J. Role of human hypoxanthine guanine phosphoribosyltransferase in activation of the antiviral agent T-705 (favipiravir). Mol. Pharmacol. 84, 615-629 (2013).
25. (a) Smither, S. J.; Eastaugh, L. S.; Steward, J. A.; Nelson, M.; Lenk, R. P.; Lever, M. S. Post-exposure efficacy of oral T-705 (favipiravir) against inhalational Ebola virus infection in a mouse model. Antiviral Res. 104, 153-155 (2014). (b) Oestereich, L.; Lüdtke, A.; Wurr, S.; Rieger, T.; Muñoz-Fontela, C.; Günther, S. Successful treatment of advanced Ebola virus infection with T-705 (favipiravir) in a small animal model. Antiviral Res. 105, 17-21 (2014).
26. Florescu, D. F.; Keck, M. A. Development of CMX001 (Brincidofovir) for the treatment of serious diseases or conditions caused by dsDNA virases. Expert Rev. Anti Infect. Ther. 12, 1171-1178 (2014).
27. Johansen, L- M.; Brannan, J. M.; Delos, S. E.; Shoemaker, C. J.; Stossel, A.; Lear, .; Hoffstrom, B. G.; Dewald, L. E.; Schornberg, K. L.; Scully, C.; Lehar, J.; Hensley, L. E.; White, J. M.; Olinger, G. G. FDA-approved selective estrogen receptor modulators inhibit Ebola virus infection. Sci. Transl. Med. 5, 190ra79 (2013).
28. Panchal, R. G.; Reid, S. P.; Tran, J. P.; Bergeron, A. A.; Wells, J.; Kota, K. P.; Aman, J.; Bavari, S. Identification of an antioxidant small-molecule with broad-spectrum antiviral activity. Antiviral Res. 93, 23-29 (2012).